Результаты усилий человека, направленные на изменение организмов в его интересах, могут служить основой для оценки действенности эволюционных факторов: мутаций, рекомбинаций и селекции. Посредством планомерного вмешательства человека в процессы изменчивости организмов "скорость эволюции" может быть значительно повышена. Таким образом, результаты планомерного разведения могут лучше, чем результаты естественно протекающих процессов показать к чему может привести длительная селекция.
Для Дарвина результаты исследования в области разведения животных и растений играли важную роль для обоснования принципа естественного отбора и реальности самой эволюции.
В этом разделе излагаются результаты исследования в области разведения живых организмов через призму микро- и макроэволюции. В связи с этим большой интерес вызывает вопрос: насколько достоверно можно по искусственной селекции (разведению) судить о естественных процессах?
На первый взгляд искусственная селекция может рассматриваться как модель ускоренного естественного отбора. Ускорение процесса изменчивости становится возможным благодаря (рис. 3.33):
Особая возможность в наиболее короткие сроки производить даже новые биологические виды предоставляется через полиплоидизацию.
В таблице 3.2 перечислены некоторые общие признаки домашних животных и культурных растений, а также ряд результатов разведения. Таблица 3.3 дает обзор целей деятельности человека по разведению животных и растений, составляющих основу этой деятельности.
Многие цели и результаты селекционной деятельности человека имеют народнохозяйственное значение, но в условиях дикой природы теряют всякий смысл. В подтверждение этого приведем следующие примеры:
Итак, селекционер отбирает мутации совсем по другим критериям, нежели природный отбор. Выведенные человеком сорта и породы, как правило, менее жизнеспособны, чем исходные формы. Большая часть выведенных экземпляров не может выжить в естественных условиях - сохранить поголовье им помогает уход со стороны человека. Животные с высокими показателями относительно слабоиммунны, т. е. подвержены заболеваниям. В случае искусственной селекции отбор происходит целенаправленно и планомерно, в то время как "естественный отбор" не предполагает наличия предварительно планирующей и управляющей инстанции.
Из этих соображений перенос искусственных взаимоотношений на естественные, природные, представляется проблематичным; общим для обеих форм является способ воздействия на генофонд: изменяются частоты аллелей, некоторые аллели подавляются, другие, напротив, получают благоприятные условия для развития Направление развития, однако, различно.
Так как с имеющимися структурами происходят многократные радикальные изменения, то границы возможностей развития резко раздвигаются. Это подтверждается следующими примерами:
Названные примеры показывают, что искусственное ускорение процессов изменчивости приводит, соответственно, и к более быстрому исчерпыванию генетического потенциала. Гораздо быстрее достигаются границы возможностей развития, потенциала изменчивости видов.
Но при этом не происходит новообразования структур. Об этом уже говорилось в разделе 3.3. Вот несколько примеров, которые должны проиллюстрировать все это еще раз:
Продолжающаяся искусственная селекция приводит к гомозиготности и, тем самым, к снижению изменчивости. Поэтому возникает термин - "тупиковый путь развития" (рис. 3.38).
Тем, что селекционер уничтожает всех измененных особей и мутантов, не соответствующих его целям, обедняется генофонд пород, подвергающихся целенаправленной селекции, при этом теряются многие аллели природных форм. Из генетического многообразия популяции выхватывается лишь одна часть. Чем меньше и однороднее генофонд популяции, тем менее разнообразны селекционные возможности и возможности развития. Таким образом, популяции попадают в генетический тупик, в котором они без опекающей руки человека либо обречены на вымирание, либо должны "выйти" из него при помощи скрещивания с природными формами (скрещивание аллелей природных форм).
Пример культурной пшеницы подходит для того, чтобы пояснить некоторые основополагающие выводы селекционного исследования. Селекционеры, занимающиеся пшеницей, вынуждены сегодня бороться с тем, что генофонд культурных сортов ржи и пшеницы для селекционных целей почти полностью исчерпан. Это означает, что популяции отдельных сортов пшеницы обладают лишь незначительным потенциалом различных аллелей. Таким образом, наступает явное сужение широты разброса генетических свойств культурной пшеницы. Селекционные возможности кажутся абсолютно исчерпанными. "Это прогрессирующее обеднение первоначального состава генов не только затрудняет выведение более устойчивых сортов, но и приводит к тому, что растения пшеницы становятся все более восприимчивыми к новым заболеваниям и неблагоприятным климатическим условиям" (FELDMAN, SEARS, 1981). Разведение лучших и более выносливых сортов пшеницы происходило за счет когда-то широко распространенного многообразия сортов пшеницы. Новые сорта, каждый из которых - "чистопородный" тип, вытеснили более примитивные сорта, составленные генетически из смеси множества различных типов. Примитивность традиционных сортов касается урожайности, а не их биологической целесообразности. Путем искусственно получаемых мутаций едва ли можно было повысить способность к изменчивости культивируемых сортов пшеницы. Дальнейшее обеднение генофонда можно, вероятно, предотвратить созданием так называемого "генного банка" имеющихся сортов пшеницы. Восстановление или даже расширение генетического потенциала можно достичь только тогда, когда будет привлечен огромный потенциал, генный резерв диких родственников наших сортов пшеницы и распространен на различные культурные формы. Итак, надежды на дальнейшее селекционное улучшение основываются на генетическом богатстве диких родственников культурных сортов пшеницы.
Первоначальным районом распространения пшеницы был Ближний и Средний Восток. На плодородных возвышенностях стран Полумесяца, которые простираются от Средиземноморского побережья на Западе вокруг сирийской пустыни до долин рек Евфрат и Тигр на Востоке, имеется особенно много диплоидных и полиплоидных видов пшеницы, обладающих замечательной способностью к изменчивости. Эта способность к изменчивости может служить богатой кладовой для селекционеров. "Дикие виды с их высокой генетической изменчивостью таят в себе множество важных в народнохозяйственном плане генов. К желательным свойствам, имеющимся в диких природных формах относятся, прежде всего, более высокое содержание белка и более высокое его качество, повышенная сопротивляемость засухе, высоким температурам, поражениям насекомыми, грибковым и вирусным заболеваниям, повышенная устойчивость ветрам и обильным выпадениям осадков, повышенная морозоустойчивость, нетребовательность к содержанию минеральных солей в почве и, наконец, более раннее созревание, Природные формы должны быть в состоянии поставлять гены, способные непосредственно повышать урожайность" (FELDMAN, SEARS, 1981).
Эти данные не оставляют сомнений, по крайней мере, в случае с пшеницей, в том, что выведенные сорта в генетическом плане сильно обеднели по сравнению с их природными предшественниками, и никоим образом не могут служить отправной точкой дальнейшего развития. Необходимость постоянно черпать из "кладовой природы" ясно показывает, что искусственная селекция, проводимая человеком, приводит не к улучшению, а к явному ухудшению сортов в плане биологической целесообразности.
Полиплоидизация
Под полиплоидизацией понимают увеличение хромосомных наборов (геномов) Если имеется три, четыре, пять, шесть или восемь наборов хромосом, то речь идет о триплоидах, тетраплоидах, цента-, гекса-, октоплоидах и т. д. Если степень плоидности выше двух, то речь идет о полиплоидах. В мире животных представлены, как правило, диплоиды (двойной набор хромосом), но нередки и гаплоиды (одинарный набор хромосом). В мире растений больше распространены полиплоиды, особенно многократно плоидны культурные растения. Например, некоторые сорта земляники тетра-, гекса- и октоплоидны. Некоторые сорта вишни имеют шестнадцать степеней плоидности.
Существует предположение, что подобные культурные сорта возникли в природе путем полиплоидизации. В действительности возможно проведение полиплоидизации экспериментальным путем посредством обработки колхицином.
Различают автополиплоидию и аллополиплоидию (рис. 3.39). При автополиплоидии (кратко - автоплоидия) умножаются хромосомы одного вида. Если, например, диплоидное растение обладает хромосомным набором АА, то автотетраплоид имеет АААА.
Между степенью плоидности и объемом ядра клеток имеется взаимосвязь: чем выше степень, тем больше объем ядра клетки, объем клетки и органов, например, фруктов, листьев, лепестков. Этим объясняется огромный размер ягоды садовой земляники по сравнению с лесной земляникой.
Аллополиплоидия происходит в случае умножения хромосомных наборов различных видов, соединенных Друг с другом путем скрещивания. Гибрид вида с хромосомным набором АА и другого вида ВВ получает набор АВ. Но гибриды в большинстве случаев стерильны. Путем полиплоидизации при помощи колхицина можно получить плодоносящий гибрид аллотетраплоидный ААВВ.
Наиболее известный пример аллоплоидного растения - культивируемая гексаплоидная мягкая пшеница (Triticum aestivum). Предполагаемый механизм ее происхождения можно проследить экспериментально. Упрощенно путь происхождения можно изобразить следующим образом (рис. 3.42). Скрещивание Triticum monococcum (культурной однозернянки, АА) с диким злаком Aegilops speltoides (ВВ) и последующая полиплоидизация дали в результате тетраплоидную двузернянку (Triticum dicoccoides, ААВВ). Скрещивание двузернянки с Aegilops squarrosa (ДД) дало яровую пшеницу (ААВВДД). При этом свою роль сыграли мутации. Другим примером является американский культивируемый хлопок (аллоплоидный гибрид американских и азиатских диких сортов), культурные сорта табака и родовой гибрид Raphanobrassica - аллотетраплоидный гибрид редьки (Ruphanus) и капусты (Brassica).
Полиплоидизация и эволюция
Автоплоидные растения не дают плодотворного скрещивания с диплоидными исходными сортами, так как у гибридов происходят нарушения мейоза. Таким образом, полиплоиды генетически изолированы от своих исходных форм. Генный обмен возможен только внутри полиплоидных и внутри диплоидных исходных форм. Тем самым при полиплоидизации в соответствии с определением биологических видов возникают новые виды. Но подобное видообразование не связано с происхождением нового генетического материала. В большей степени происходит "сбор" имеющегося генетического материала различных видов. Возникающие при этом виды обладают часто измененными свойствами и признаками по сравнению с исходными видами, а не принципиально новыми органами. Таким образом, видообразование путем полиплоидизации однозначно следует отнести к области микроэволюции. Поэтому об "эволюции культурной пшеницы" можно говорить только в плане микроэволюции.
Заключение
|
Табл. 3.2. Примеры целенаправленной селекции. |
|
Наиболее часто встречающиеся признаки культурных растений: |
|
|
Признаки домашних животных: |
|
|
Другие примеры из мира животных и растений: |
|
|
Табл. 3.3. Основные направления селекционной деятельности человека |
|
|
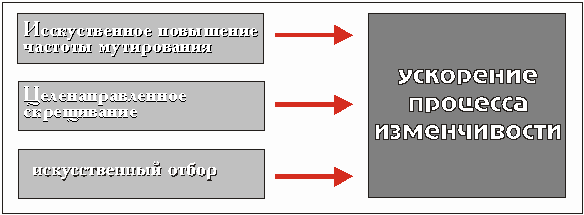
Рис. 3.33 Повышение скорости процесса изменчивости (микроэволюции) путем планомерных селекционных мероприятий.

3.34 Пестролистность (Galathea)

Рис. 3.35. Махровые лепестки у выведенной формы цветка маргаритки (внизу). Новая форма отличается от природной (снимок наверху) большим количеством лепестков и более высокой интенсивностью окраски. Окраску красного цвета имеют теперь уже и природные формы.
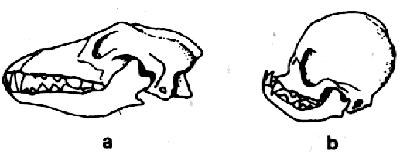
Рис. 3.36 Воздействие искусственного отбора у собачьих. а – волк (подвергшийся мутации прапредок всех собачьих пород), в – пекинес с ярко выраженным укороченным черепом (По Meyer, Daumer 1981 масштаб различен).

Рис.3.37. Из сизого голубя (посередине) в течении немногих сотен лет был выведен целый ряд пород голубей (рисунок Lotti Haller).
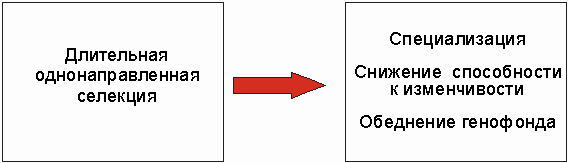
Рис.3.38 Длительная однонаправленная естественная и искусственная селекция приводит к узкой специализации, снижению способности к изменчивости и обеднению генофонда. Этим часто характеризуется и разнонаправленная селекция.
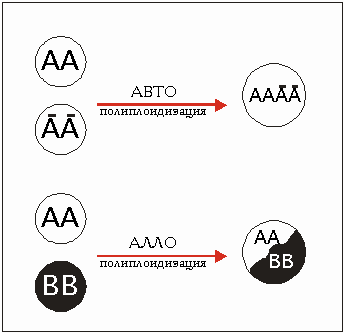
Рис. 3.39. Авто- (а) а аллополиплоидизация (b); a - набор генов двух индивидуумов одного вида, b - набор генов двух индивидуумов различных видов.
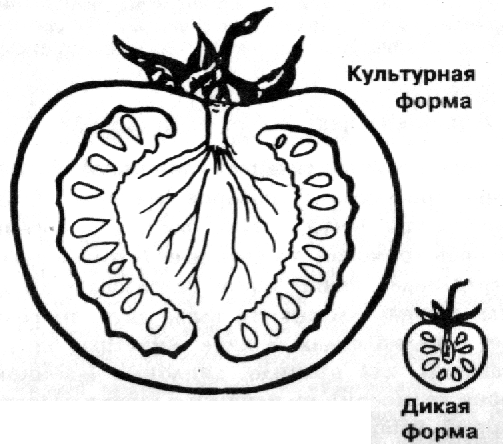
Рас. 3.40. Гигантский рост у томатов. (По Osche, 1972.)
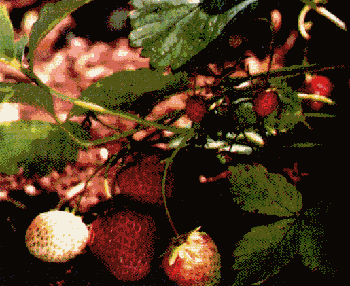
Рас. 3.41. Одна из основных форм и культурная форма земляники. Значительное увеличение размеров ягод у культурной формы происходит зачастую за счет потери вкусовых качеств.

Рис. 3.42. Предположительное происхождение культурной
пшеницы. Произошло двойное скрещивание видов (АА+ВВ - > АВ, ААВВ+ДД - >
АВД,) с последующей полиплоидизацией (АВ - ААВВ, АВД - >AАВВДД). А, В и Д
символизируют гаплоидные хромосомные наборы. Внес ли свой "вклад" в развитие
дикого злака В-ген - это вопрос спорный. (DEIHL, 1980, OSCHE.)