Для возникновения более высоких систематических категорий согласно эволюции требуется гораздо больше времени. Бактерии и в этом случае кажутся наиболее подходящими на роль первопроходцев. Приобретение нового генного материала относится к основным проблемам при создании новых конструкций. У бактерий относительно легко можно было бы проверить появление новых, ранее даже в прототипе не встречавшихся ферментов. Для получения таких эволюционных новинок рассматривается два способа: удвоение генного материала внутри генома с последующим превращением и получение генетического материала от других организмов.
Удвоение (дупликация)
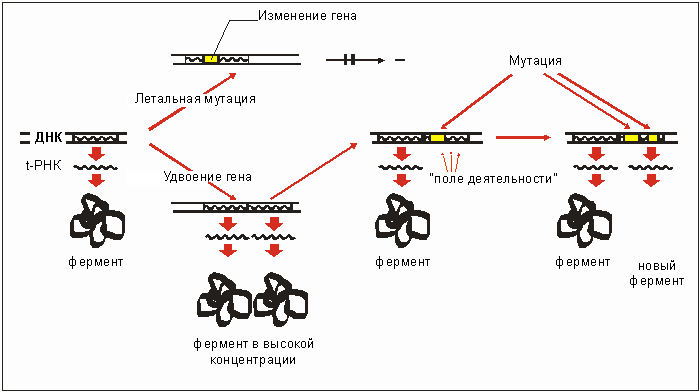
После удвоения ген остается неизменным, в то время как второй возникший в результате удвоения ген служит "полем деятельности эволюции" для мутаций и тем самым для апробации новых функций (рис. 3.31). У бактерий наблюдается умножение генов, например, таких, ферменты которых участвуют в дезактивировании антибиотиков (рис. 3.26). В другом случае умножается ген дефектной синтетазы (фермент, соединяющий молекулы), которая обуславливает лишь незначительное увеличение каких-либо аминокислот в белке. В обоих случаях приумножение гена позволяет настолько поднять уровень концентрации фермента, что обеспечивается полная выживаемость. В одном случае становится возможной более быстрое дезактивирование, в другом - жизненно важный протеиновый синтез.
Посредством удвоения удается предотвратить потерю генной функции, оно может быть сколь угодно велико и наступать спонтанно с частотой до 10-3. Однако такое умножение гена нестабильно и исчезает, как только ослабевает воздействие селекции. Какие условия должны быть выполнены для того, чтобы в умноженном гене вследствие мутаций возникла абсолютно новая энзиматическая (ферментная) функция, вы узнаете в разделе 4.3.
Векторный перенос генов
Если организмы должны стать абсолютно новыми системами, насколько .что позволяют новые условия жизни, то достичь это возможно через передачу генов векторами, как в случае с плазмидами и бактериофагами.
Это можно продемонстрировать на примере: плазмида, обладающая интактным лактогеном (ген, расщепляющий молочный сахар), переносится на лак-отрицательный штамм Е. coli (штамм, особи которого не могут использовать лактозу), при этом происходит превращение принимающих клеток в лак-положительный штамм. Штамм опять приобретает способность использовать лактозу в качестве источника С.
Однако посредством подобной передачи генов не возникает штаммов с новой генетической информацией, а лишь происходит новое перераспределение уже имеющихся генов (этот процесс сравним с процессом рекомбинации, см. раздел 3.3.2).
Результаты генного картирования
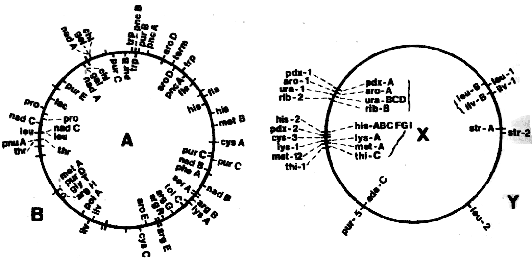
Возможности генного картирования и рядности ДНК привели к сравнительному изучению последовательности генов в хромосомах бактерий. При этом оказалось, что родственные штаммы имеют похожие, а неродственные - абсолютно различные генные карты (рис. 3.32). Генные карты энтеробактерий Е. coli К 12 и Salmonella typhimurium LT 2 очень похожи. Это же подходит и для Streptomyces coelicolor и Nocardia mediterranea. Сравнение генных карт актиномицет с картами энтеробактерий не выявляет никакой схожести. Сравнение отражает абсолютно разные конструкции, прежде всего в биохимическом и физиологическом аспектах. Между этими группами никогда не наблюдалось генных обменов какого-либо типа. Явственно проглядывается тенденция ограничить генный обмен рамками одного генотипа. Горизонтальный перенос генов, посредством конъюгации и трансформации бактерий из различных таксономических групп, например, семейств и родов, поставил бы под вопрос усилия по систематизации бактерий. Однако, как учит опыт, перенос, в действительности, не имеет значения для макроэволюции.
Иммунитет против одинаковых и родственных плазмид и защита при помощи ферментов, ограничивающих их воздействие, отличаются тем, что они, расщепляя ДНК чужих видов, гарантируют стабильность штаммов бактерий. Поэтому созданные усилиями генной инженерии новые штаммы могут доставить определенные трудности.
Заключение
Рис. 3.31. Гипотеза удвоения. После удвоения гена (дубликации) клетка может подвергнуться любым изменениям посредством имеющегося теперь уже в двойном размере гена. Там могут наслаиваться любые мутации (поле деятельности эволюции). Таким образом, в результате ряда мутации, может возникнуть новый фермент. (По KAMPFE, 1980,)
Рис. 3.32. Две группы геномов: энтеробактерии (А - Escherichia
coli; В - Salmonella typhimurium) u актиномицеты (X - Streptomyces coelicolor,
Y - Nocardia mediterranea). Величины геномов N. mediterranea и S.coelicolor приведены
к одному масштабу. Порядок расположения объединенных скобками генотипов неизвестен.
(По SANDERSON, 1976 и SCHUPP, 1975.)