В наиболее широком смысле можно все изменения наследственности обозначать как мутации. При этом не имеет значения, где они происходят - в соматических клетках или в гаметах. Для эволюции важны лишь мутации, меняющие наследственность эмбриональных клеток, так как именно они будут унаследованы следующим поколением. Те мутации, которые вызывают изменение фенотипа (это внешняя форма проявлений организма, тогда как генотип - совокупность наследуемых факторов), были известны уже очень давно. Но лишь с тех пор как была открыта структура ДНК, стали понятными некоторые молекулярные механизмы.
В первую очередь следует назвать генные мутации: в этом случае изменения затрагивают краеугольный камень ДНК. Соответствующий триплет может теперь кодировать другую аминокислоту, что приводит к изменению первичной структуры соответствующего протеина (рис. 4.5). Это может, но не обязательно должно иметь своим следствием более или менее глубокие изменения функции протеина. А упомянутая выше точечная мутация при определенных условиях может сделать протеин абсолютно неспособным функционировать (рис. 4.6). Примером этому может служить уже упомянутая серповидноклеточная анемия (раздел 3.3.3). При этом заболевании в бетагемоглобиновой цепочке человека заменена одна лишь аминокислота. Если этот ген является гетерозиготным (одна мутированная и одна нормальная аллель), болезнь не очень ярко выражена (ее распознают, кроме всего прочего, по тому, что красные кровяные тельца при определенных условиях принимают серповидную форму). Если ген является все же гомозиготным (две мутированные аллели), то следствием этого может быть тяжелая форма анемии (малокровие). Как раз человеческий гемоглобин очень хорошо исследован, и определены соответствующие аминокислотные цепочки многих тысяч людей. При этом до сегодняшнего дня описано свыше трех сотен вариантов гемоглобина человека, и это, конечно же, малая часть от существующих в действительности форм. Точечные мутации. однако, не обязательно должны приводить к изменению аминокислотных цепочек, так K.IK различные триплеты иногда кодируют одинаковые аминокислоты (вырожденность генетического кода, см. рис. 6.10). Подобные мутации называются также "молчащими" и могут быть выявлены лишь в результате анализа цепочки ДНК. С другой стороны, может получиться так, что в ходе точечной мутации будет заменена одна аминокислота без наступления функциональных изменений: тогда речь идет о нейтральной мутации (см. раздел 3.6.2). Если на функцию оказывается негативное влияние, как, например, в случае гомозиготной формы серповидноклеточной анемии, то перед нами предстает мутант с отрицательным селективным значением. Имеется множество позиций аминокислот, которые могут подвергнуться каким-либо изменениям. Такие позиции называются инвариантными. В зависимости от функции протеина речь может вестись о большем (например, гистоновые белки) или меньшем (например, фибринопептиды) число позиций. Если функция в связи с наступлением новых условий окружающей среды улучшается (см. резистентность по отношению к антибиотикам, раздел 3.3.3), то можно говорить о положительном значении отбери. Но точечная мутация происходит в той части ДНК, где закодированы регуляторные функции. И хотя структура протеина там не изменится, подобные мутации могут иметь, вероятно, огромное значение для фенотипа индивидуума.
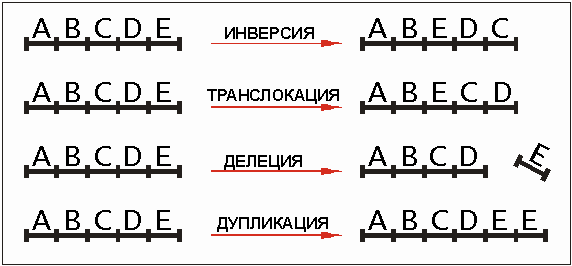
Из классической генетики известны хромосомные мутации (рис. 4.7). К ним относятся: делеция (выпадение части хромосомы), инверсия (поворот участка хромосомы на 180 градусов), транслокация (ген перемещается с одного места хромосомы на другое) или дупликация генов. Эти изменения касаются не только целых хромосомных отрезков. Возможно "выпадение" генных фрагментов и отдельных генов, они также могут быть удвоены или "прыгнуть" на другое место. Особенно у бактерий, но также и у более высокоразвитых живых существ встречается генный обмен между индивидуумами (см. об этом в разделе . 3.3.4). Генная дупликация, наряду с точечной мутацией, рассматривается как один из самых важных для эволюции типов мутаций, т. к. в этих случаях происходит увеличение генетического материала. Посредством точечной мутации удвоенный ген может быть инактивирован на место, он тогда не будет переведен в аминокислотную цепочку. Некоторые из этих генов называются псевдогенами. Изменения, соответствующие хромосомным мутациям, кажется, имеются и в области отдельных генов. Таким образом, может произойти так, что в результате делеции один или несколько нуклеотидов выпадут, будут транспортированы на другое место и введены тем самым в ген. Мутировать могут даже "старт-" и "стоп-сигналы" на ДНК и почти что "консервировать" ген. Бывает, что в результате мутации удваиваются целые хромосомы.
В качестве примера следует привести трисомию по 21 паре у человека - это наличие дополнительной хромосомы (болезнь Дауна). Умножаться могут не только хромосомы, но и целые хромосомные блоки, что играет особо важную роль в выведении полезных растений. Биологи, сторонники эволюционного учения, сходятся, однако, во мнении, что подобные мутации генных композиций при возникновении новых конструкций не играли сколько-нибудь значительной роли. Таким образом, не создается новой информации (несущей новый смысл цепочки ДНК), но лишь увеличивается уже имеющаяся. По всей видимости, при образовании новых видов играет определенную роль слияние различных хромосом. Так, например, у различных популяций мышей, изолированных на фермах в Северной Америке, выявлено различие в числе хромосом.
Подводя итог, можно констатировать, что все поддающиеся экспериментальному доказательству явления мутации внутри основных типов происходят в рамках микроэволюционных процессов (см. главу 3 и 10). Следует заметить, что это не объяснение, а лишь чисто эмпирическое заключение. Для эволюционно-теоретических рассуждений очень важно знать нормы мутаций. На основании многочисленных наблюдений сегодня можно уже назвать довольно точные цифры. При этом доля жизнеспособных мутантов не во всех случаях одинакова. Было установлено, что в определенных областях мутации происходят особенно часто, но еще точно не известно, отчего это происходит. Вероятно, в этих случаях отключаются механизмы восстановления или репарации ДНК.
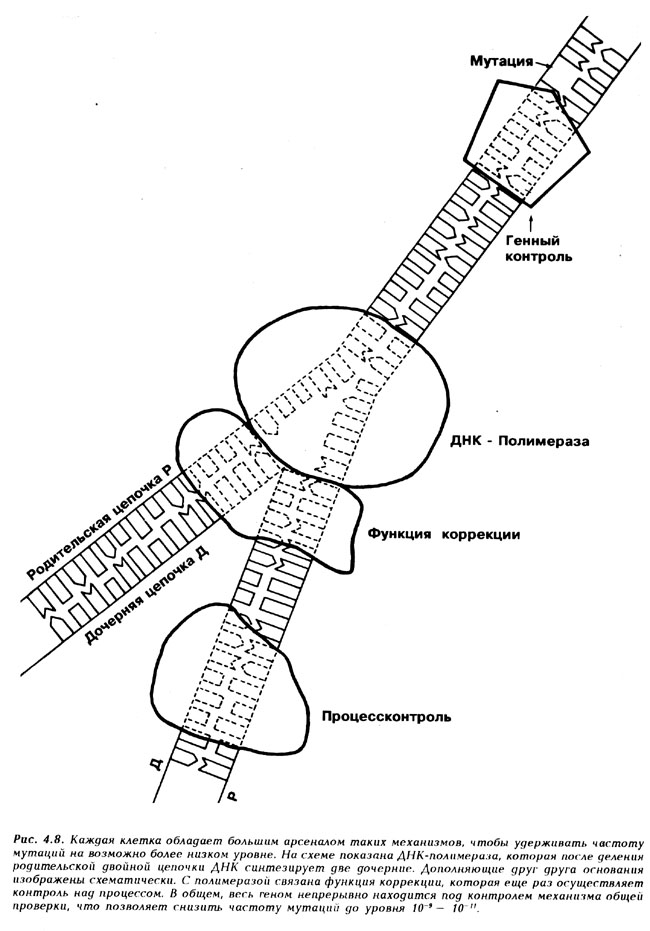
Следует дифференцировать два понятия: "частота мутаций" и "норма мутаций". Под "частотой мутаций" мы понимаем вероятность, с которой при транскрипции ДНК происходит неверный синтез нуклеотида, при этом не имеет значения, приводит ли это к биологической смерти индивидуума, или к селективному преимуществу. "Частота мутаций" - это понятие, относящееся к нуклеотидам и принципиально отличающееся от понятия "нормы мутаций". Под последним понимают определенную установленную в ходе наблюдений процентную долю индивидуумов популяции, которые проявили за период жизни одного поколения мутацию по одному из признаков. Таким образом, нормы мутаций относятся к признакам, подвергнувшимся мутации (это может соответствовать одному или нескольким генам, а также распространяться на нуклеотид). Взаимосвязь между этими двумя понятиями сложна и не совсем ясна, т. к., во-первых, не всякий обмен оснований приводит к изменению признаков (например "молчащая" мутация); во-вторых, результаты многих обменов оснований являются смертоносными для индивидуумов, то есть совсем не обязательно, что они проявятся в потомстве. Кроме того, изменение различных оснований одного гена может, вероятно, привести к аналогичному изменению признака. Все клетки настроены на то, чтобы сдерживать мутационную частотность на возможно более низком уровне. Для этой цели они располагают целым каскадом механизмов контроля и восстановления (рис. 4.8). Если правильная последовательность "основных кирпичиков" ДНК зависела бы только от комплементарности (взаимного соответствия) пар оснований, то тогда на сто сопряжении нуклеотидов приходилось бы от 1 до 10 ошибок. Частота мутаций применительно к нуклеотидам составляла бы 10-1 - 10-2. Благодаря объемной структуре своего активного центра ДНК-полимераза допускает уровень ошибок до 10-4 - 10-5. Дополнительно ДНК-полимераза осуществляет функцию коррекции, благодаря которой распознает неправильный нуклеотид и удаляет его. Тем самым, возможность ошибок снижается с 10-6 до 10-8. За ДНК-полимеразой следует еще одна процедура контроля, которая сверяет дочернюю цепочку с родительской и при необходимости коррегирует ее, что уменьшает вероятность ошибки до 10-9 10-10 . Эти цифры настолько малы, что только сравнение может помочь их понять. Эта книга сдержит по приблизительным оценкам 750000 букв. И если бы секретарша могла работать с такой же точностью, как ДНК-синтетаза какой-либо бактерии, то она, сделав 3500 копий этой книги, в общей сложности допустила бы лишь одну ошибку. При этом она имела бы право работать над каждой копией не более трех минут!
Эти механизмы препятствуют тому, чтобы при удвоении ДНК происходили мутации, но наряду с этим клетки обладают еще и механизмом восстановления, который препятствует распространению в наследственном материале мутаций под влиянием радиации или химических веществ. Ясно видно, что усилия, которые затрачивает клетка на то, чтобы сдерживать частоту мутаций на низком уровне, чрезмерно велики. Мутации, как правило, вредны для организма. Поэтому абсолютно непонятно, как "примитивная" клетка должна была вообще выжить без механизмов контроля и восстановления (см. главу 5).
Что же касается такого явления, как антитела (это протеины, которые служат для защиты от бактерий), то для них могла быть намеренно повышена норма мутаций, чтобы увеличить многообразие этих антител. Вероятно, это имеет некоторые селективные преимущества. В общем, мутации происходят, но при этом (в данном случае речь тоже идет о вероятности) норма поднимается с 10-5 до 10-11. Под этим подразумевается следующее. Среди сотни тысяч бактерий, например, найдется в среднем один индивидуум, который несет в себе мутацию только одного признака, что уже считается высокой нормой мутаций. Однако, в культуре бактерий присутствует до миллиарда клеток, так что вероятность найти в популяции один индивидуум с одной определенной мутацией может быть достаточно высокой. Но, как уже было сказано, от гена к гену это чрезвычайно разнится. Например, вряд ли может такое случиться, чтобы нашелся индивидуум, несущий мутацию в гистон-гене (гистоны - это протеины, которые связаны с ДНК и могут выполнить свою задачу только в совокупности с крайне специфической цепочкой аминокислот, они регулируют биосинтез и-РНК). Вообще, мутации можно наблюдать настолько часто, что каждый человек несет в своих зародышевых клетках и в клетках тела большое число мутаций. Некоторые биологи придерживаются даже мнения, что процесс старения связан с частотой мутаций в клетках организма. Все сказанное выше подкрепляет мысль, что мутации являются не целенаправленными, а случайными явлениями. Поэтому если более последовательно принимать во внимание ограничивающие условия (особенно отбор), то можно описать мутационные процессы с помощью статистических закономерностей.
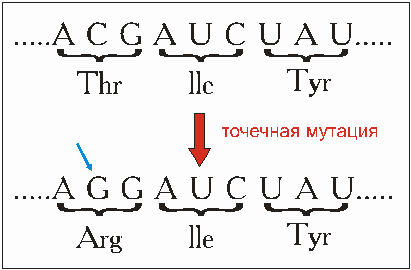
Рис. 4.5. Точечная мутация изменяет последовательность нуклеотидов в ДНК ч, соответственно, в и-РНК (здесь показано, что в и-РНК имеете основания тимина стоит основание урацил). Это может привести, как в данном примере, к изменению последовательности аминокислот протеина и к возможному изменению его функции. Благодаря обмену основания цитозина на гуанин этот триплет не кодирует более аминокислоту треонин, а кодирует аргинин. lle обозначает аминокислоту изолецин, Tyr - аминокислоту тирозин. Накоплением подобных точечных мутации объясняется в общеупотребительной теории эволюции большинство эволюционных процессов. Подтверждается ли это экспериментами, будет рассматриваться в разделе 4.3.
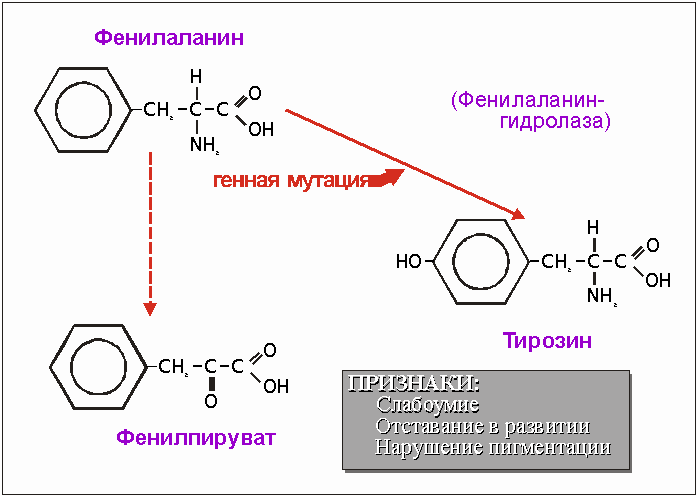
Рис. 4.6. В результате генной мутации у некоторых людей нарушаются образование фермента фенилаланингидролазы. Это приводит к тому, что аминокислота тирозин не может больше образовываться, а из фенилаланина образуется фенилпируват, который появляется в моче. Одна мутация гена ведет к. целому ряду серьезных последствий. К счастью, сегодня эта наследственная болезнь (фенилкетонурия) излечивается.
Рис. 4.7. Четыре важных хромосомных мутации. Буквы означают в этом случае видимые в световой микроскоп фрагменты хромосом, которые могут быть перестроены. Подобные механизмы можно наблюдать и в генах, и в генетических фрагментах. Особо важным для эволюции считается генная дупликация (удвоение), которая даже через световой микроскоп различима как изменения последовательности генов в хромосомах.
Рис. 4.8. Каждая клетка обладает большим арсеналом таких механизмов, чтобы удерживать частоту мутаций на возможно более низком уровне. На схеме показана ДНК-полимераза, которая после деления родительской двойной цепочки ДНК синтезирует две дочерние. Дополняющие друг друга основания изображены схематически. С полимеразой связана функция коррекции, которая еще риз осуществляет контроль над процессом. В общем, весь геном непрерывно находится под контролем механизма oбщей проверки, что позволяет снизить частоту мутации до уровня 10-9 - 10-11 .