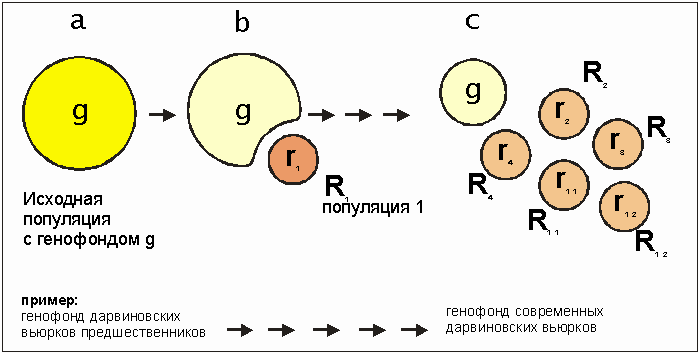
Каковы последствия продолжающегося разделения видов на отдельные частичные популяции? Конкретнее: как изменяется генофонд популяции при многократно повторяющихся разделениях популяции? Приведенный ниже рис. 3.57 дает изображение рассматриваемых впоследствии процессов: Если от популяции А отделяется часть (рис. 3.57) генофонд r1 части популяции R, получает, как правило, только одну часть ассортимента аллелей изначального общего генофонда g. Редкие аллели могут, к примеру, для частей популяций просто пропасть. В лучшем случае часть популяции получает равное с исходной число различных аллелей.
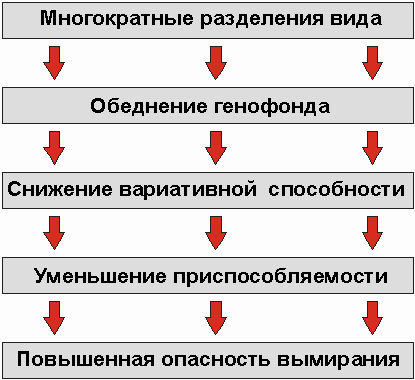
Посредством генного сдвига отделенная часть популяции может развиться в новую популяцию. Это развитие может даже привести к возникновению нового вида. Если и дальше часть популяции R2 подвергается разделению, то генофонды частей обычно имеют другой состав, нежели генофонд r2. Путем различных микроэволюционных процессов субпопуляция R2 может настолько отдалиться от субпопуляции R1, что обе более не могут плодотворно скрещиваться друг с другом. При этом может сохраниться "мостик" (возможность скрещиваться), связывающий с исходной популяцией (сравните с Rana pipiens в разделе 10.4). Отделенная часть популяции соответствующей породы или вида R может и дальше разделяться на частичные популяции R11 и R12, имеющие генофонды r11 и r12, и т. д. с соответствующими последствиями, как и при первом разделении. Генофонды отделенных пород становятся все более и более обедненными относительно исходж,1х пород, т. е. они обладают все меньшим количеством различных аллелей. Тем самым сокращается число комбинаций аллелей частях популяций по сравнению с исходной: возможности рекомбинаций сужены. Это означает не что иное, как то, что разделенные породы или виды обладают меньшей изменчивостью. Это зависит от количества различных аллелей и их возможных комбинаций. Из этого опять-таки следует, что на изменение окружающей среды они могут реагировать менее гибко: то есть перед нами пониженная приспособляемость. И при изменении условий окружающей среды породы с обедненным генофондом размножаются хуже, чем исходный вид со всем своим генетическим богатством. Поэтому им в наибольшей степени грозит опасность вымирания.
Голландский зоолог Duyvene de Wit хорошо описал эти процессы (Цит. по KAHLB, 1984, с. 87. ср. с рис 3.58):
"Если пограничная популяция прокладывает путь к новому жизненному пространству, она не может взять все гены материнской популяции, но лишь часть из них. Каждый подвид (или вид), выходящий из предыдущего, становится обладателем еще более бедного генофонда. Следовательно, потеря основы генофонда есть та цена, которую должен платить подвид или вид за преимущественное право прийти в этот мир. Если же процесс видообразования многократно повторяется, то, в конце концов, появляются такие виды, чьи генофонды настолько опустошены, что бывает достаточно относительно небольшого изменения в условиях окружающей среды, чтобы вызвать их вымирание. Пониженная сопротивляемость условиям окружающей среды как результат пониженной способности к рекомбинации приводят, в конце концов, к некому минимальному генетическому уровню, за гранью которого выживание невозможно. Трагическая судьба чрезвычайно узко специализированных видов или пород поэтому и приводит к непременной генетической смерти."
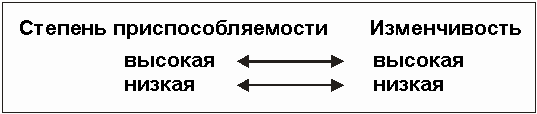
Отделенные популяции могли бы, конечно, благодаря воздействию отбора быть лучше приспособлены к их специфической окружающей среде, чем были бы приспособлены исходные популяции, однако, быть лучше приспособленным означает вместе с тем быть уже приспособленным (см. рис. 3.59). "Быть лучше приспособленным" - это понятие идет рука об руку с понятием крайне высокой (экстремальной) степени специализации, пусть даже и за счет богатства генофонда. Приспособляемость и специализация, с одной стороны, и развитие от низших форм к высшим, с другой, - суть две разные вещи. Это доказывает существование неспособных летать насекомых на островах подверженных сильным ветрам, или пещерных животных (см. раздел 3.3.3).
Об эволюционной стадии можно вести речь только в смысле микроэволюции.
Можно ли обедненный в результате разделения видов генофонд полностью восстановить путем мутаций и рекомбинаций (см. рис. 3.6), эту проблему невозможно решить теоретическими рассуждениями. Вместо этого пусть говорит красноречивый пример: гепард, самый быстрый на свете зверь, несмотря на выдающиеся способности спринтера и охотника, оказался на грани вымирания Признаком этого является большая подверженность заболеваниям и высокая доля аномальных сперматозоидов. Следует заметить также, что после охоты он настолько ослаблен, что не в состоянии защищать свои добычу от конкурентов (львов, леопардов, гиен). Обширные исследования вскрыли вероятную причину угрозы для гепарда: популяция гепардов не проявляет почти никакой изменчивости. Это означает, что один гепард почти полностью похож на другого. Суженая изменчивость приводит к генетическому единообразию. Более точно это могло быть подтверждено сравнением анализов крови и иммунологическими исследованиями. Угроза вымирания гепардов, которая охватила большую часть их первоначальной области обитания, очевидно, является результатом такого генетического единообразия, так как это связано со снижением возможности приспосабливаться к условиям окружающей среды.
Каким же образом этот вид потерял свою генетическую изменчивость? Наиболее убедительная гипотеза исходит из того, что когда-то в прошлом этот вид понес огромные потери: популяция прошла (и, возможно, многократно) "через игольное ушко". Тем самым генетический состав вида потерял многие аллели. Популяция, понесшая большие потери и едва избежавшая вымирания, вынуждена была в дальнейшем расти путем близкородственного скрещивания, что и должно было привести к генетическому единообразию. О возможных причинах этих масштабных потерь в популяции можно рассуждать лишь на философском уровне. Все то же самое можно наблюдать и у искусственно выведенных форм, что демонстрирует пример популяции гепардов и что явственно следует из приведенных выше рассуждений слишком высокая степень приспособленности связана со слишком сильным обеднением генофонда и сильно ограниченной способностью выживания.
Заключение
Рис 3.57. Происходящие одни за другими отделение популяции ( R1, R2, и т. д.) г генофондами, r1 и r2 и т.д. от исходной популяции с генофондом g. а - исходная ситуация; b - ставшая самостоятельной часть популяции R с генофондом r1, с - дальнейшее отделение популяции, ставших подвидами R2, R3 и R4 генофондами r2 , r3 u r4. Популяция R1 еще раз разделилась на части R11 и R12 с генофондами r11 и r12. Дарвиновские вьюрки (см. рис. 3.2) представляют, очевидно, стадию с, насчитывающую 14 видов, не полностью изолированных друг от друга в плане скрещияания, для сравнения см. раздел 3.2
Рис 3.58. Наиболее часто встречающаяся взаимосвязь между продолжающимся разделением вида и опасностью вымирания.
Рис. 3.59. Взаимосвязь между степенью приспособляемости и вариативностью.
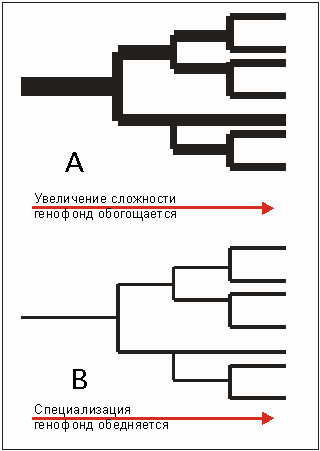
Рис. 3.60. Две противоположные схемы восприятия последствий происходящего деления видов. Наверху - восприятие эволюционной теории, внизу - учения о сотворении.